Cornea epithelium is subject to constant trauma and shedding of the surface epithelium, and replenishment is from epithelial cells beneath and peripheral to the central desquamating epithelium. The origin of the corneal epithelium appears to reside in the Palisades of Vogt (PoV), where a population of immortal stem cells resides, characterized by possessing enormous potential for clonogenic cell division. These cells, like other stem cells in the body, have inherent properties which enable them to accomplish error-free replication which avoids development of abnormal differentiation and cellular dysfunction; a low mitotic rate and asymmetrical DNA segregation are essential in this error-free proliferation of these relatively primitive, poorly differentiated cells.
Limbal stem cells lack keratin 3, a differentiation marker of the corneal epithelium, but express keratins such as K14, K15, and K19. Cell kinetic studies show that limbal stem cells and their progeny, known as transient amplifying cells (TACs), remain within the proliferative compartment, where they actively divide before eventually committing to terminal differentiation. These cells (stem cells and TAC) are located exclusively in the limbal region, with the stem cells in the basal epithelium and TAC occurring in the basal and suprabasal levels, extending up to the superficial layers. The corneal epithelium is then maintained by cellular proliferation of these cells which then migrate further onto the central cornea and become terminally differentiated. Terminal differentiation of corneal limbal stem cells is controlled by a combination of microenvironmental cues, molecular signals, and intrinsic transcriptional programs. The lack of a vascular supply and the presence of special elements within the corneal basement membrane, and differences in vitamin A concentrations may be some of the primary factors responsible for the terminal differentiation events.
The absence or dysfunction of corneal stem cells leads to loss of corneal epithelial proliferative capacity, resulting in conjunctivalization of the corneal surface or, in severe cases, persistent epithelial defects with neovascularization and scarring. Such disorders include primary dysfunction such as aniridia or secondary to destruction of limbal stem cells, either traumatically (e.g., alkali burns) or immunologically (e.g., Stevens Johnson syndrome).
It is well recognized that corneal transplantation alone is insufficient for achieving vision restoration and corneal healing in these cases; however, limbal stem cells transplantation can restore the corneal epithelium. It is important to note that limbal stem cell transplantation is unlikely to provide a definitive solution unless the underlying pathology—particularly chronic ocular surface irritation from conditions such as sicca syndrome, meibomian gland dysfunction, lagophthalmos, trichiasis, distichiasis, keratinized posterior lid margin, or immune-mediated ocular surface diseases—is corrected. Thus, limbal stem cell transplantation may set the stage for the ultimate visual rehabilitation step (i.e., keratoplasty.
Our experience with limbal stem cell transplantation has been quite satisfactory. The majority of our patients have advanced ocular surface diseases, most commonly associated with cicatricial pemphigoid or severe atopy and less frequently with Stevens–Johnson syndrome.
New techniques of limbal stem cell transplant include direct limbal cell transplantation (DLCT), cultivated limbal epithelial transplantation (CLET), and mesenchymal stem cell transplantation (MSCT). The field has been rapidly evolving in recent years, with the goal of developing customized therapies tailored to different severities of limbal stem cell deficiency. Efforts have focused on understanding limbal stem cell regulation, expanding cell sources for transplantation, improving cultivation methods, and refining limbal stem cell transplantation techniques.
References
 |
|
|
|
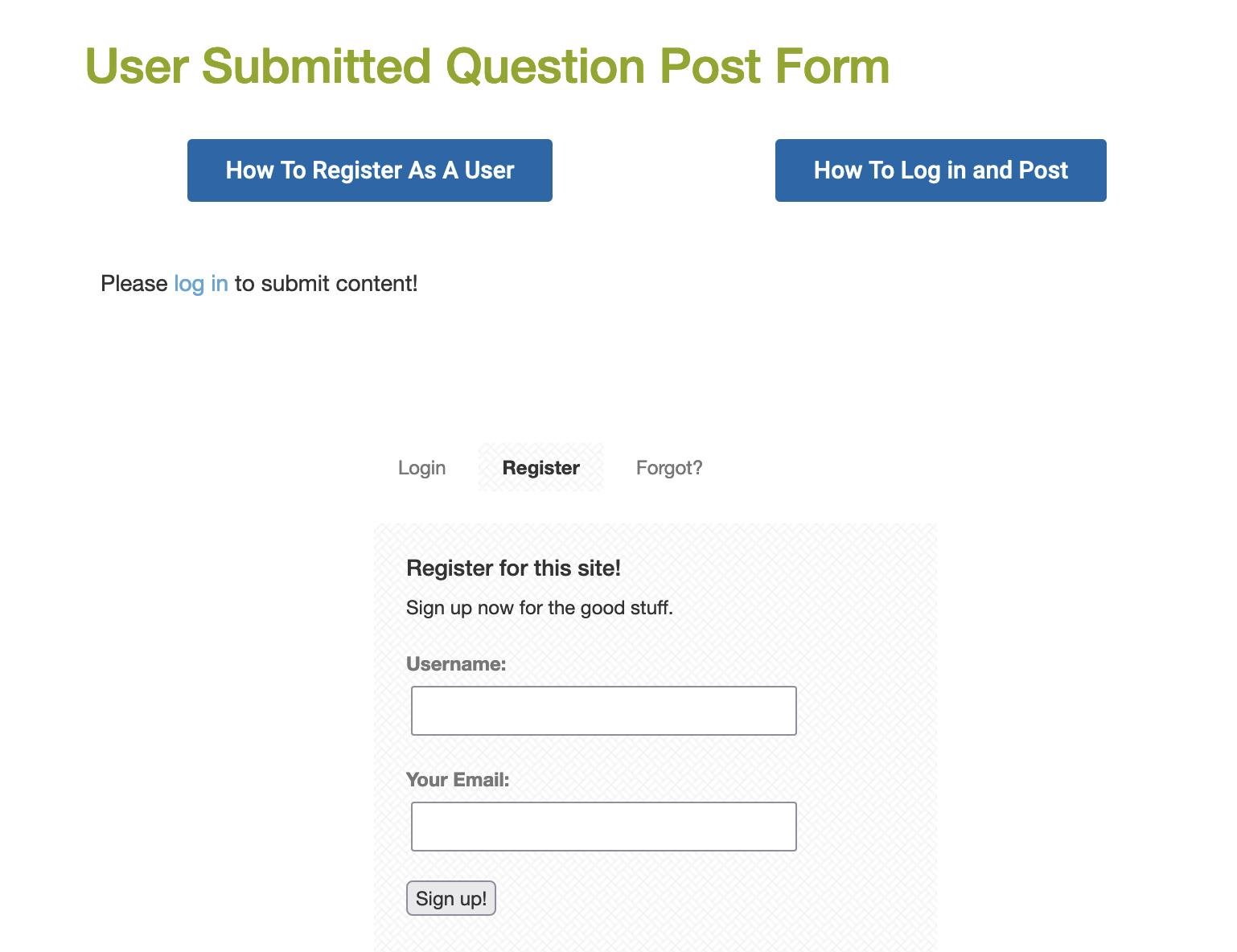
Click on the Register link
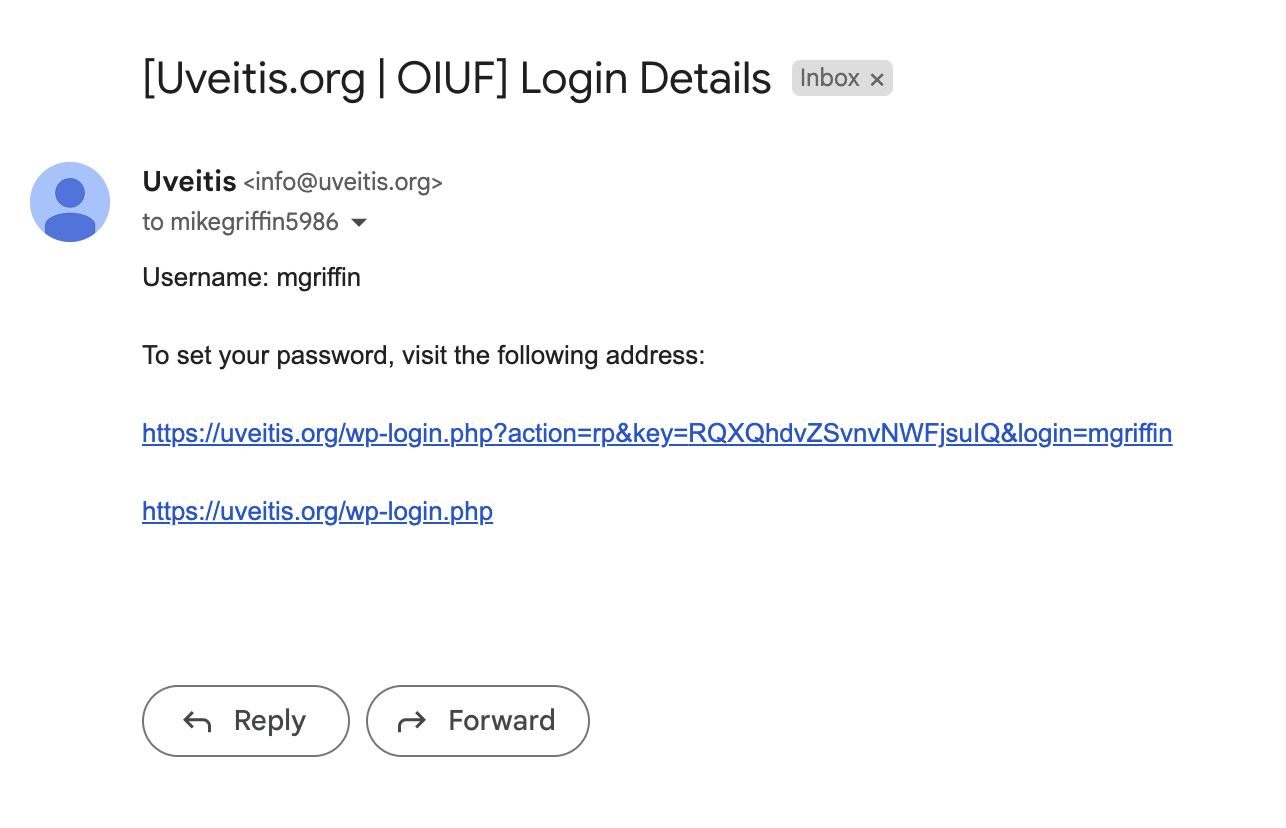
In the email you receive, click on the change password link.
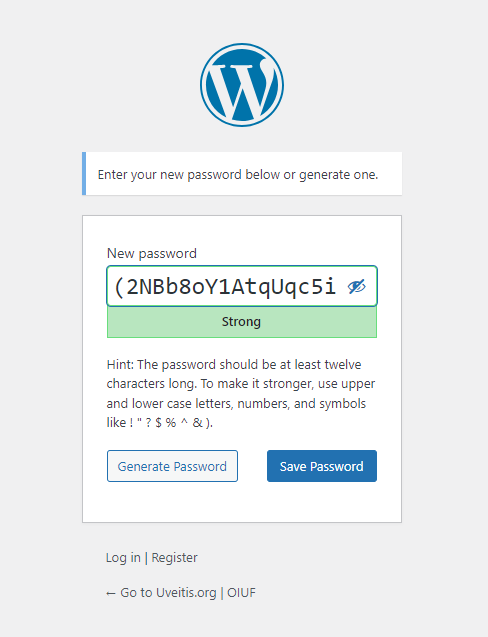
Change your password